РАЗДЕЛ 11. ГОРМОНАЛЬНАЯ РЕГУЛЯЦИЯ ОБМЕНА ВЕЩЕСТВ И ФУНКЦИЙ ОРГАНИЗМА
VII. Регуляция обмена ионов кальция и фосфатов
В организме взрослого человека содержится в среднем 1000 г кальция, из которых 99% находится в костях в виде кристаллов гидроксиапатита [Са10(РО4)6(ОН)2Н2О]. Остальные 1% кальция представлен в плазме крови, поступая из кишечника и костной ткани в процессе резорбции.
Кальций выполняет не только структурную функцию в костях, но и играет важную роль в мышечном сокращении, проницаемости клеточных мембран для ионов калия, натриевой проводимости, секреции гормонов и свёртывании крови. Концентрация кальция в клетках зависит от его уровня во внеклеточной жидкости. Нормальные пределы концентрации Са2+ в плазме крови здоровых людей составляют 2,12 — 2,6 ммоль/л (9 — 11 мг/дт). Кальций в плазме представлен в следующих формах:
- несвязанного, ионизированного кальция (около 50%);
- ионов кальция, связанных с белками, в основном альбумином (45%);
- недиссоциирующих комплексов с цитратом, сульфатом, фосфатом и карбонатом (5%).
Ионизированный кальций является биологически активной фракцией, его концентрация поддерживается в пределах 1,1 — 1,3 ммоль/л. Изменение уровня кальция может нарушить многие процессы, включая возбудимость нервных и мышечных клеток, работу кальциевого насоса и гормональную регуляцию метаболизма. Концентрация Са2+ в плазме регулируется с высокой точностью: изменение всего на 1% активирует гомеостатические механизмы, восстанавливающие равновесие.
Основные регуляторы обмена Са2+ в крови — паратгормон, кальцитриол и кальцитонин.
А. Паратгормон
Паратгормон (ПТГ) — полипептид из 84 аминокислот, который повышает уровень ионов кальция и снижает уровень фосфатов в плазме крови.
1. Синтез и секреция ПТГ
ПТГ синтезируется в паращитовидных железах как предшественник — препрогормон, содержащий 115 аминокислот. В эндоплазматическом ретикулуме отщепляется сигнальный пептид, и образуется прогормон, который превращается в зрелый гормон (ПТГ, 1-84) в аппарате Гольджи. Паратгормон хранится в секреторных гранулах и может расщепляться на активные фрагменты. Секреция ПТГ регулируется уровнем ионов кальция в плазме: гормон выделяется при снижении концентрации кальция.
2. Роль паратгормона в регуляции обмена кальция и фосфатов
Органы-мишени для ПТГ — кости и почки. В клетках этих органов находятся специфические рецепторы, взаимодействующие с паратгормоном, что инициирует каскад событий, приводящий к активации аденилатциклазы и увеличению концентрации цАМФ. Это, в свою очередь, способствует мобилизации ионов кальция из внутриклеточных запасов.
 Читайте также:
Читайте также:
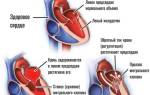
В костной ткани рецепторы ПТГ расположены на остеобластах и остеоцитах. Связываясь с ними, паратгормон стимулирует секрецию инсулиноподобного фактора роста 1 и цитокинов, что активирует остеокласты и приводит к распаду костного матрикса, высвобождая Са2+ и фосфаты.
В почках ПТГ увеличивает реабсорбцию кальция в дистальных канальцах, снижая его экскрецию с мочой, и уменьшает реабсорбцию фосфатов. Паратгормон также стимулирует синтез кальцитриола, который усиливает всасывание кальция в кишечнике.
3. Гиперпаратиреоз
Вторичный гиперпаратиреоз возникает при хронической почечной недостаточности и дефиците витамина D3, что приводит к гипокальциемии из-за нарушения всасывания кальция. Секреция паратгормона увеличивается, но не может нормализовать уровень кальция. Это может привести к остеопорозу. Третичный гиперпаратиреоз возникает при автономной гиперсекреции паратгормона.
4. Гипопаратиреоз
Гипопаратиреоз, вызванный недостаточностью паращитовидных желез, приводит к гипокальциемии, что может вызвать неврологические и сердечно-сосудистые нарушения, а также спазмы.
Б. Кальцитриол
Кальцитриол, синтезируемый из холестерола, повышает уровень кальция в плазме крови.
1. Строение и синтез кальцитриола
- Читайте также:
В коже 7-дегидрохолестерол превращается в холекальциферол под воздействием УФ-излучения. Затем холекальциферол гидроксилируется в печени и почках, образуя кальцитриол, активную форму витамина D3. Синтез кальцитриола стимулируется паратгормоном и низкой концентрацией фосфатов и ионов кальция.
2. Механизм действия кальцитриола
Кальцитриол воздействует на тонкий кишечник, почки и кости, связываясь с внутриклеточными рецепторами и индуцируя транскрипцию генов, ответственных за синтез белков, которые обеспечивают всасывание кальция и фосфатов. В почках он увеличивает реабсорбцию этих ионов. Недостаток кальцитриола может привести к рахиту и остеомаляции.
3. Рахит
Рахит — заболевание, связанное с недостаточной минерализацией костной ткани, вызванное дефицитом кальция и витамина D3. Основные способы профилактики — правильное питание и достаточная инсоляция.
В. Роль кальцитонина в регуляции обмена кальция
Кальцитонин — полипептид, секретируемый парафолликулярными клетками щитовидной железы, который снижает уровень кальция в крови, ингибируя активность остеокластов и реабсорбцию кальция в почках. Секреция кальцитонина зависит от уровня эстрогенов.
Паратгормон — полипептидный гормон, регулирующий обмен кальция и фосфора, повышая уровень кальция и снижая уровень фосфатов в крови. Он активирует резорбцию костной ткани и влияет на кишечник, усиливая всасывание кальция.
Секреция паратгормона регулируется концентрацией ионизированного кальция в крови. При гиперпаратиреозе наблюдается нарушение фосфорно-кальциевого обмена, а при гипопаратиреозе — тетания и другие нарушения.
- Читайте также:
Библиография: Булатов А. А. Паратгормон и кальцитонин, в кн.: Биохимия гормонов и гормональной регуляции, под ред. Н. А. Юдаева, М., 1976; Машковский М. Д. Лекарственные средства, М., 1977; Романенко В. Д. Физиология кальциевого обмена, Киев, 1975; Руководство по клинической эндокринологии, под ред. В. Г. Баранова, Д., 1977; Стуккей А. Л. Околощитовидные железы, в кн.: Физиология эндокринной системы, под ред. В. Г. Баранова, Д., 1979.
Действие паратгормона на почки
В дистальном нефроне находятся рецепторы паратгормона и кальциевые рецепторы. Это позволяет внеклеточному Са++ оказывать прямое воздействие через кальциевые рецепторы и опосредованное — через модификацию уровня паратгормона в крови, влияя на почечный кальциевый гомеостаз. Внутриклеточным медиатором действия паратгормона является ц-АМФ, уровень которого в моче служит биохимическим маркером активности околощитовидных желез. Почечные эффекты паратгормона включают:
- Увеличение реабсорбции Са++ в дистальных канальцах. При избыточном выделении паратгормона экскреция Са++ с мочой возрастает из-за увеличения фильтрации кальция при гиперкальциемии.
- Увеличение экскреции фосфата. Паратгормон ингибирует Na-зависимый транспорт фосфата в проксимальных и дистальных канальцах.
- Увеличение экскреции бикарбоната из-за угнетения его реабсорбции в проксимальных канальцах, что приводит к ощелачиванию мочи. При избыточной секреции паратгормона это может вызывать тубулярный ацидоз из-за интенсивного выведения щелочного аниона.
- Увеличение клиренса свободной воды и объема мочи.
- Увеличение активности витамин D-la-гидроксилазы, которая синтезирует активную форму витамина D3, способствующую всасыванию кальция в кишечнике и влияющую на обмен кальция.
Таким образом, при первичном гиперпаратиреозе избыточное действие паратгормона проявляется в виде гиперкальцийурии, гипофосфатемии, гиперхлоремического ацидоза, полиурии, полидипсии и увеличенной экскреции нефрогенной фракции цАМФ.
Действие паратгормона на кости
Паратгормон оказывает как анаболический, так и катаболический эффект на костную ткань. Эти эффекты можно разделить на две фазы: в ранней фазе происходит мобилизация кальция из костей для быстрого восстановления баланса с внеклеточной жидкостью, а в поздней фазе стимулируется синтез костных ферментов, таких как лизосомальные энзимы, которые способствуют резорбции и ремоделированию костной ткани. Основным объектом действия паратгормона в костях являются остеобласты, так как остеокласты, по-видимому, не имеют его рецепторов. Под воздействием паратгормона остеобласты вырабатывают различные медиаторы, среди которых выделяются провоспалительный цитокин интерлейкин-6 и фактор дифференцировки остеокластов, которые стимулируют дифференциацию и пролиферацию остеокластов. Остеобласты также могут подавлять функцию остеокластов, вырабатывая остеопротегерин. Таким образом, резорбция костей остеокластами стимулируется опосредованно через остеобласты. При этом увеличивается высвобождение щелочной фосфатазы и экскреция с мочой гидроксипролина — маркера разрушения костного матрикса.
Двойственное действие паратгормона на костную ткань было открыто в 30-е годы XX века, когда установили его резорбтивное и анаболическое влияние. Однако лишь 50 лет спустя, на основе исследований с рекомбинантным паратгормоном, стало известно, что длительное постоянное воздействие избытка паратгормона вызывает остеорезорбцию, а пульсовое интермиттирующее введение стимулирует ремоделирование костной ткани. В настоящее время только синтетический паратгормон (терипаратид) имеет лечебный эффект при остеопорозе, а не просто приостанавливает его прогрессирование, и разрешен к применению FDA США.
Действие паратгормона на кишечник
Пратгормон не влияет напрямую на абсорбцию кальция в желудочно-кишечном тракте. Его эффекты осуществляются через регуляцию синтеза активной формы витамина D (1,25(OH)2D3) в почках.
Другие эффекты паратгормона
В опытах in vitro были выявлены дополнительные эффекты паратгормона, физиологическая роль которых еще не до конца ясна. Например, установлено, что он может изменять кровоток в кишечных сосудах, усиливать липолиз в адипоцитах и увеличивать глюконеогенез в печени и почках.
Витамин D3, упомянутый ранее, является вторым важным гуморальным агентом в регуляции кальциевого гомеостаза. Его действие, направленное на повышение всасывания кальция в кишечнике и увеличение концентрации Са++ в крови, оправдывает название «гормон D». Биосинтез витамина D — это сложный многоэтапный процесс. В крови человека могут одновременно находиться около 30 метаболитов, производных или предшественников наиболее активной формы 1,25(ОН)2-дигидроксилированного витамина. Первым этапом синтеза является гидроксилирование углеродного атома в положении 25 стирольного кольца витамина D, который поступает с пищей (эргокальциферол) или образуется в коже под воздействием ультрафиолетовых лучей (холекальциферол). На втором этапе происходит повторное гидроксилирование молекулы в положении 1а специфическим ферментом проксимальных почечных канальцев — витамин D-1а-гидроксилазой. Из множества производных витамина D только три обладают выраженной метаболической активностью: 24,25(OH)2D3, 1,24,25(OH)3D3 и 1,25(OH)2D3, при этом только последний действует направленно и в 100 раз сильнее остальных. Витамин D, взаимодействуя со специфическими рецепторами в ядре энтероцитов, стимулирует синтез транспортного белка, который переносит кальций и фосфат через клеточные мембраны в кровь. Обратная отрицательная связь между концентрацией 1,25(ОН)2 витамина D и активностью 1а-гидроксилазы обеспечивает ауторегуляцию, предотвращая избыток активного витамина D.
Витамин D также оказывает умеренный остеорезорбтивный эффект, который проявляется только в присутствии паратгормона. Он также тормозит синтез паратгормона околощитовидными железами в дозозависимом порядке.
- Читайте также:
Кальцитонин является третьим основным компонентом гормональной регуляции обмена кальция, однако его действие значительно слабее, чем у предыдущих двух агентов. Кальцитонин — это 32-аминокислотный белок, который секретируется парафолликулярными С-клетками щитовидной железы в ответ на повышение концентрации внеклеточного Са++. Его гипокальциемическое действие осуществляется за счет ингибирования активности остеокластов и увеличения экскреции кальция с мочой. Физиологическая роль кальцитонина у человека до сих пор не установлена, так как его влияние на кальциевый обмен незначительно и перекрывается другими механизмами. Полное отсутствие кальцитонина после тотальной тиреоидэктомии не вызывает физиологических отклонений и не требует заместительной терапии. Значительный избыток этого гормона, например, у больных медуллярным раком щитовидной железы, не приводит к серьезным нарушениям кальциевого гомеостаза.
Регуляция секреции паратгормона в норме
Основным регулятором секреции паратгормона является внеклеточный кальций. Даже небольшое снижение концентрации Са++ в крови вызывает мгновенное увеличение секреции паратгормона. Этот процесс зависит от степени и продолжительности гипокальциемии. Первичное кратковременное снижение концентрации Са++ приводит к высвобождению накопленного в секреторных гранулах паратгормона в течение первых нескольких секунд. Через 15-30 минут гипокальциемии увеличивается также синтез паратгормона. Если стимул продолжается, то в течение первых 3-12 часов (у крыс) наблюдается умеренное повышение концентрации матричной РНК гена паратгормона. Длительная гипокальциемия стимулирует гипертрофию и пролиферацию паратиреоцитов, что можно обнаружить через несколько дней или недель.
Кальций воздействует на околощитовидные железы и другие органы через специфические кальциевые рецепторы. В 1991 году Brown впервые предположил существование таких структур, а позже рецептор был выделен, клонирован и изучены его функции и распределение. Это первый из рецепторов у человека, который распознает непосредственно ион, а не органическую молекулу.
Человеческий Са++-рецептор кодируется геном на хромосоме 3q13-21 и состоит из 1078 аминокислот. Молекула рецептора включает большой N-концевой внеклеточный участок, центральное мембранное ядро и короткий C-концевой внутрицитоплазматический хвост.
Открытие рецептора объясняет происхождение семейной гипокальциурической гиперкальциемии, при которой обнаружено более 30 различных мутаций гена рецептора. Также недавно установлены активирующие мутации Са++-рецептора, приводящие к семейному гипопаратиреозу.
Са++-рецептор широко экспрессирован в организме, не только в органах, участвующих в метаболизме кальция (околощитовидные железы, почки, С-клетки щитовидной железы, клетки костной ткани), но и в других органах (гипофиз, плацента, кератиноциты, молочные железы, гастрин-секретирующие клетки).
Недавно был обнаружен другой мембранный кальциевый рецептор на паратиреоцитах, плаценте и проксимальных почечных канальцах, роль которого требует дальнейшего изучения.
Среди других модуляторов секреции паратгормона следует отметить магний. Ионизированный магний влияет на секрецию паратгормона, но менее выраженно, чем кальций. Высокий уровень Mg++ в крови (например, при почечной недостаточности) угнетает секрецию паратгормона. В то же время гипомагнезиемия приводит не к увеличению, а к парадоксальному снижению секреции паратгормона, что связано с угнетением его синтеза при недостатке ионов магния.
Витамин D также влияет на синтез паратгормона через генетические транскрипционные механизмы. 1,25-(ОН) D подавляет секрецию паратгормона при низком уровне кальция в сыворотке и увеличивает внутриклеточную деградацию его молекулы.
Другие гормоны оказывают модулирующее действие на синтез и секрецию паратгормона. Например, катехоламины, действуя через β-адренергические рецепторы, усиливают секрецию паратгормона, особенно при гипокальциемии. Антагонисты β-адренорецепторов обычно снижают уровень паратгормона в крови, но при гиперпаратиреозе этот эффект минимален из-за изменения чувствительности паратиреоцитов.
Глюкокортикоиды, эстрогены и прогестерон стимулируют секрецию паратгормона. Эстрогены могут также модулировать чувствительность паратиреоцитов к Са++, стимулируя транскрипцию гена паратгормона и его синтез.
Секреция паратгормона регулируется ритмом его высвобождения в кровь. Установлено, что помимо стабильной тонической секреции существует пульсовая, составляющая 25% от общего объема. При остром возникновении гипокальциемии или гиперкальциемии сначала реагирует пульсовая секреция, а затем, через 30 минут, и тоническая.

